
![]()
![]()
El límite de cada célula está determinado por la membrana plasmática mediante la cual se relaciona con el ambiente y responde al mismo. Además, en la célula eucarionte, varios organelos están formados por membranas cuya composición y estructura son como las de la membrana plasmática.
La existencia de lípidos parcialmente solubles en agua propicio el origen de las membranas que han posibilitado la formación de un sistema abierto que permite el intercambio de materia y energía, no solo con el medio externo, sino entre los diferentes compartimientos que tienen las células eucariontes.
A la flexibilidad y resistencia que proporcionan los lípidos de la membrana se añade la especificidad que poseen todas las células debido a la presencia de proteínas y carbohidratos que, dada su variabilidad química, proporcionan la base para efectuar las múltiples funciones que caracterizan a los sistemas vivos.
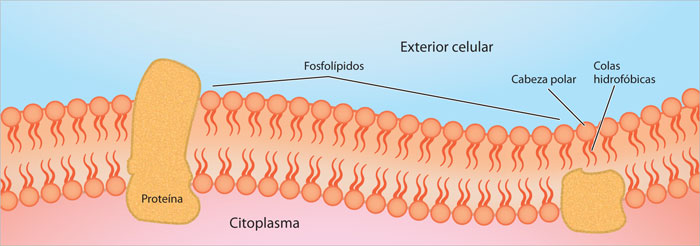
Los componentes químicos de las membranas biológicas son lípidos variados, proteínas y oligosacáridos y están arreglados en forma de un mosaico fluido.
Debido a que los lípidos de membrana tienen una zona polar hidrofílica y una parte hidrofóbica, forman bicapas en un ambiente acuoso con dos propiedades importantes: impiden el paso de las sustancias polares a través de la membrana, por lo que estas moléculas y los iones deben transportarse mediante proteínas especificas; además la bicapa es muy estable, flexible y resistente debido a las interacciones no covalentes que se establecen entre sus moléculas —como las fuerzas de Van der Waals e interacciones hidrofóbicas.
La bicapa de lípidos es fluida y su consistencia es similar a la del aceite de oliva. Los lípidos que la forman pueden moverse lateralmente, pero rara vez un lípido de una de las caras de la bicapa pasa a la otra. Las membranas son muy sensibles a los cambios de temperatura del medio; una temperatura baja provoca la gelificación de las membranas, mientras que una temperatura alta produce el efecto contrario. Ambas situaciones impiden que las membranas desarrollen sus funciones óptimamente; sin embargo, muchos organismos son capaces de regular la fluidez de la membrana mediante el cambio en la composición de sus lípidos.
Los distintos tipos celulares tienen en sus membranas plasmáticas y en las membranas de los organelos, una composición variada de lípidos; esto indica la estrecha relación de estas sustancias con las funciones específicas que desempeñan. Por ejemplo, cierto tipo de lípidos se concentra en algunas zonas del retículo endoplásmico y del aparato de Golgi para formar sus curvaturas características.
Si bien los lípidos de membrana representan la estructura básica de las membranas celulares, las proteínas que se encuentran en ellas desempeñan funciones específicas muy importantes. La mayoría de las membranas plasmáticas consta de 50% de lípidos y 50% de proteínas, pero esta proporción varia de acuerdo con el tipo de membrana analizado. La coexistencia de una gran variedad de lípidos y proteínas, que presentan, en general, gran movilidad, permitió que en 1972, Jonathan Singer y Garth Nicolson propusieran el nombre de mosaico fluido para la estructura de las membranas celulares.
Singer y Nicolson reconocieron dos tipos de proteínas de membrana: las periféricas, que pueden separarse fácilmente de la capa lipídica y las integrales, que se encuentran insertadas total o parcialmente en la bicapa. Muchas de las proteínas integrales atraviesan totalmente las membranas exponiendo sus extremos en ambos lados de la membrana; las diferentes regiones de estas proteínas (dominios) desempeñan, a menudo, funciones diversas: la región expuesta hacia el exterior puede ser la zona de reconocimiento de una hormona; la región transmembranal ancla la proteína a la membrana; la región citosólica puede interactuar con proteínas diversas para desencadenar reacciones que se traducirán en una acción especifica, por ejemplo, la salida de glucosa de una célula hepática.
Las proteínas de la membrana plasmática pueden ser, por ejemplo, enzimas como la adenilil ciclasa que permite la formación del cAMP (adenosín monofosfato cíclico) a partir de ATP o las proteínas transportadoras de electrones, que intervienen en los procesos de oxidorreducción relacionados con la producción de ATP y que se encuentran en las membranas de las mitocondrias, de los cloroplastos y de los procariontes.
El transporte de la mayoría de las sustancias a través de las membranas se hace por medio de proteínas transportadoras o acarreadoras; por canales, que son pasajes hidrofílicos a través de los cuales pueden pasar principalmente pequeños iones inorgánicos, y mediante las denominadas "bombas" que mueven sustancias en contra de un gradiente de concentración; es decir, de un medio menos concentrado de la sustancia a un medio más concentrado de la misma.
Además de permitir el paso de sustancias de uno a otro lado de las membranas celulares, las proteínas pueden actuar como receptoras de señales químicas como hormonas, factores de crecimiento o neurotransmisores; estos interactúan con las proteínas receptoras, que mediante un cambio de conformación desencadenan una gran variedad de respuestas intracelulares. Las proteínas de membrana se encuentran también involucradas en la comunicación y adhesión entre células vecinas, que es una condición para la multicelularidad, asunto que se tratara más adelante.
Otra importante función de las proteínas membranales está relacionada con el ciclo de exocitosis- endocitosis; con el tráfico de sustancias que se lleva a cabo entre el retículo endoplásmico, el aparato de Golgi y las vesículas relacionadas con ellos; las proteínas de membrana tienen también importantes vínculos con las sustancias que constituyen la matriz extracelular y con el citoesqueleto.
La importancia de la correcta conformación y localización de las proteínas de membrana, se pone de manifiesto cuando el gen que las codifica sufre alguna mutación. En la fibrosis quística, una mutación en el gen que corresponde a la proteína que permite la salida de Cl- hacia la luz del tracto respiratorio, impide que este ion, junto con el Na+, formen un ambiente ligeramente hipertónico que propicie la salida de agua para hidratar el moco que recubre el epitelio respiratorio. Los resultados de este trastorno son el desarrollo de infecciones bacterianas y un intercambio de gases que se dificulta por la permanencia del moco espeso que es difícil de eliminar.
En la mayoría de las proteínas transmembranales y en algunos lípidos se presentan carbohidratos constituidos por pocas unidades de monosacáridos (denominados oligosacáridos) en la superficie expuesta hacia el exterior de la célula. Estos azucares se agregan generalmente en el retículo endoplásmico rugoso y en el aparato de Golgi. Las glicoproteínas y los glicolípidos son muy abundantes en la membrana plasmática, y en muchas células animales, constituyen una capa denominada glicocálix, muy desarrollada sobre todo en la superficie de las células epiteliales del intestino. Los carbohidratos de la membrana juegan un importante papel en el reconocimiento celular, en la reacción antigeno-anticuerpo y en la adhesión celular necesaria para la formación de tejidos.
La entrada y salida de sustancias de la célula y su tráfico interno difieren de acuerdo con su naturaleza fisicoquímica: una hormona esteroidea podrá pasar a través de la membrana fosfolipídica debido a su naturaleza no polar, mientras las sustancias polares serán incapaces de atravesarla. Es, por lo tanto, imprescindible la presencia de proteínas transportadoras y canales que faciliten el movimiento de los iones y moléculas en ambas direcciones; asimismo es necesario contar con una fuente de energía que posibilite su movimiento cuando fluyan en contra de un gradiente de concentración, es decir, desde el sitio donde la concentración de la sustancia es menor, hacia donde es mayor.
Existen tres tipos de transporte de sustancias a través de membranas. Por un lado, la difusión simple y la difusión facilitada, que reciben el nombre genérico de transporte pasivo, el cual propicia el movimiento de sustancias a favor de un gradiente de concentración. Por otro lado, el transporte activo que, como ya se mencionó, utiliza energía en forma de ATP para transportar sustancias o que aprovecha la entrada de un ion a favor de su gradiente de concentración, para introducir, por ejemplo, glucosa o un aminoácido en contra de su gradiente.
Difusión simple| La difusión simple de las sustancias a través de la membrana se encuentra restringida a gases como oxígeno, bióxido de carbono, moléculas hidrofóbicas y pequeñas moléculas polares sin carga eléctrica como el agua y el etanol.
El movimiento de los solutos que se desplazan por difusión simple, siempre se hace a favor de un gradiente de concentración, es decir, van desde el sitio de mayor concentración del soluto hacia el sitio de menor concentración. En este proceso no se involucran las proteínas.
La ósmosis es un caso especial de transporte de agua. El agua tiende a atravesar la membrana de las células dependiendo de la diferencia de concentración de solutos. Una solución que tenga mayor concentración de solutos que el interior de la célula, se dice que es hipertónica; mientras que si la solución tiene una concentración más baja de solutos que la célula se dice que es hipotónica. El movimiento de agua siempre se hará desde una solución hipotónica a una hipertónica. Si la célula se coloca en agua destilada, el agua se difundirá al interior de la célula, provocando un aumento de volumen y finalmente se reventará; pero si la célula se pone en una solución saturada de solutos se encogerá.
Las células del cuerpo humano normalmente están en estado isotónico, es decir, en ambos lados de la membrana se encuentra la misma concentración de solutos; por lo tanto, no hay una tendencia clara del agua para entrar o salir. Sin embargo, de acuerdo con las condiciones metabólicas de las células, éstas pueden en un momento dado ser hipertónicas o hipotónicas con respecto al medio. En el primer caso, para restaurar la isotonía, la célula permite la salida de iones, para que se iguale la concentración celular de solutos con el exterior y, si la célula es hipotónica con respecto al medio, la isotonía se recupera con el ingreso de iones.
En las células de plantas, algas, hongos y bacterias que poseen una pared celular y que están en un ambiente hipotónico, el ingreso del agua no les provocará ruptura. En una planta, la entrada constante de agua permite su acumulación en la vacuola central; ésta ejerce una presión sobre la pared celular manteniendo la planta completamente hidratada o turgente; pero si el medio es isotónico, la planta se marchita debido a que el agua no tiende a entrar o salir de la célula.
Los organismos que carecen de paredes celulares y que viven en medios hipertónicos o hipotónicos presentan mecanismos especiales de regulación osmótica. Los protozoarios, como Paramecium caudatum, que habitan en agua dulce, tienen unas vacuolas especiales llamadas pulsátiles o contráctiles que recogen el agua excedente, tan rápido como entra en la célula.
Difusión facilitada | Es el mecanismo más simple por el cual moléculas como los azúcares, aminoácidos y diversos iones, se mueven a favor de un gradiente de concentración, en ambos sentidos de la membrana celular. Las proteínas de membrana que facilitan este tipo de transporte, reciben el nombre genérico de canales y acarreadores. Los más estudiados son los canales iónicos que transportan iones de carga positiva como Na+, K+ y Ca2+ , que participan en el cambio de los potenciales de membrana, importantes en la conducción nerviosa o en la exocitosis de vesículas que transportan neurotransmisores u hormonas como la insulina.
Las proteínas acarreadoras (carriers), llamadas también transportadoras o, en algunas ocasiones permeasas, presentan un cambio de conformación cuando la molécula que debe ser transportada se pone en contacto con ellas. El cambio de forma del acarreador permite la transferencia de la molécula al interior de la célula, sin que exista un canal propiamente dicho. El acarreador más conocido es el que permite la entrada de la glucosa al interior del eritrocito. La concentración de glucosa en el plasma sanguíneo es más elevada que el interior del eritrocito, por lo que la glucosa entra a favor de un gradiente de concentración.
Los canales proteicos son pasajes formados por aminoácidos hidrofílicos que permiten el paso de pequeños iones sin necesidad de que exista un cambio de conformación de la proteína. En general, los canales son muy selectivos y, como en su mayoría conducen pequeños iones, se les conoce como canales iónicos. Algunos canales de iones permanecen abiertos todo el tiempo, mientras otros, los llamados canales con compuerta, sólo se abren en respuesta a una señal química, física, mecánica o eléctrica.
Transporte activo| El transporte activo requiere la presencia de las denominadas "bombas" activadas por ATP o ATPasas; la energía del ATP se usa para mover iones y moléculas en contra de un gradiente de concentración químico o eléctrico. Este tipo de transporte desempeña un papel importante en las células y en los organelos: 1] permite la introducción de nutrientes al interior de la células, aunque éstos se encuentren en una concentración más baja que en el citosol; 2] asegura la expulsión de los productos de secreción y de desecho hacia el medio, y 3] finalmente, permite el mantenimiento de la concentración de iones como el Na+, K+ y Ca2+.
Las ATPasas más conocidas son la "bomba" de sodio y potasio que se encuentra en casi todas las células animales y la H+ ATPasa, responsable de la acidificación del jugo gástrico. Éstos son ejemplos de transporte activo directo, porque se agregan o se quitan fosfatos a partir del ATP de ciertas regiones de la ATPasa, ocasionando un cambio de conformación que propicia la entrada o salida de las sustancias.
Las proteínas involucradas en el transporte activo indirecto no utilizan ATP. El ejemplo más conocido es el del cotransporte de sodio/glucosa: la proteína transportadora recibe el sodio que penetra en la célula a favor de un gradiente de concentración. El sodio se une primeramente a una región de la proteína transportadora, provocando un cambio en su conformación, lo que propicia la entrada de la glucosa que tiene una concentración más baja en el exterior. En este caso, la entrada de sodio favorece el ingreso de la glucosa en contra de un gradiente de concentración. El transporte activo puede ser uniporta, cuando sólo una sustancia se transporte hacia adentro o hacia afuera; sinporta, cuando dos sustancias se transportan en la misma dirección; y antiporta, cuando ambas se mueven en dirección opuesta.