
![]()
![]()
Además de los organelos que ocupan una parte importante del volumen de la célula eucarionte, existe otro nivel de organización que consiste en una red de proteínas que se encuentra en el núcleo y en gran parte de los espacios que dejan entre sí los organelos en el citosol. Se trata del citoesqueleto, una estructura flexible y muy dinámica, cuya primera función es proveer un armazón que da forma a los distintos tipos celulares de los eucariontes. El citoesqueleto también forma estructuras relacionadas con distintas funciones como el desplazamiento de las células, el movimiento interno de estructuras como los cromosomas o el transporte de vesículas de neurotransmisores que van desde el cuerpo celular de una neurona hasta el sitio de sinapsis.
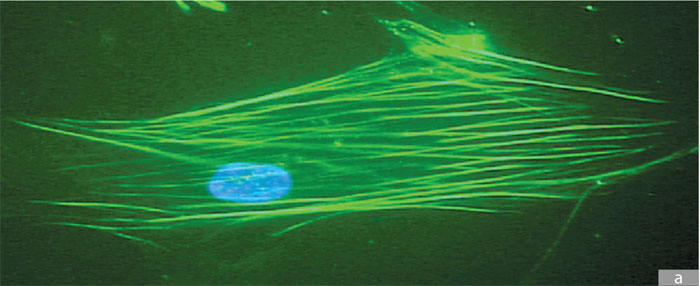
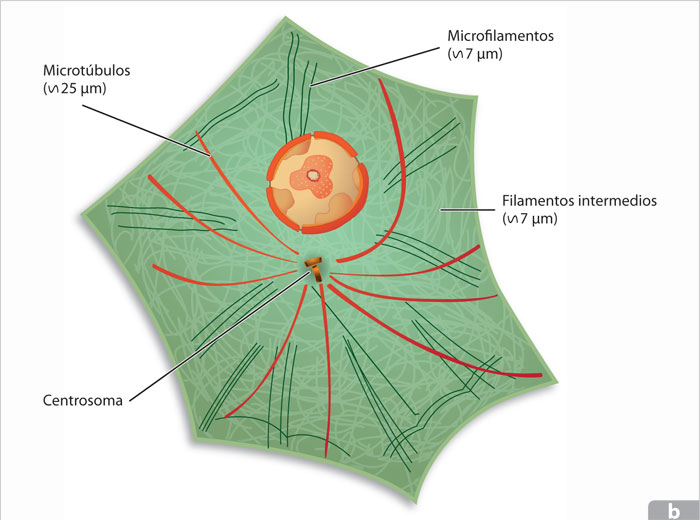
El citoesqueleto está compuesto por tres tipos de de filamentos, perfectamente distinguibles por su composición proteica y por su diámetro: los microfilamentos, constituidos por dos filamentos entrelazados, formados por la unión de unidades de actina, miden de 6 a 8 μm y son los más delgados; los microtúbulos, con un diámetro de 24 μm, están formados por subunidades de la proteína tubulina; y los filamentos intermedios que miden 10 μm. Estos últimos, a diferencia de los dos primeros, están constituidos por distintos tipos de proteínas, dependiendo del tejido donde se encuentren; así, por ejemplo, en el núcleo encontramos la laminina, en las células epiteliales queratina y en las células musculares desmina.
En la mayoría de las células eucariontes encontramos los tres representantes del citoesqueleto, aunque en sitios diferentes dependiendo de la función que desempeñan; por ejemplo, las microvellosidades de las células del epitelio intestinal están sostenidas por filamentos de actina; mientras los desmosomas, que mantienen unidas a estas células, se encuentran unidos por una red de queratinas; por último, los microtúbulos se encuentran en estrecha relación con organelos como el retículo endoplásmico y el aparato de Golgi.
En la organización y localización de los organelos membranosos, los microtúbulos participan activamente; por ejemplo, cuando los microtúbulos se desorganizan con la presencia de ciertos compuestos, el retículo endoplásmico pierde su organización básica. Los microtúbulos se organizan a partir del centrosoma o centro de organización de los microtúbulos, que en las células animales generalmente tiene un par de centriolos. En los hongos y en las plantas, el centrosoma carece de centriolos y lo mismo ocurre con algunas células animales, como los cigotos recién fecundados. Los microtúbulos también forman el aparato mitótico que permite la distribución adecuada de los cromosomas durante la mitosis y meiosis.
Los cilios y flagelos de los eucariones, al igual que los centriolos y cuerpos basales, están compuestos por microtúbulos. Su estructura es esencialmente la misma: los cilios y flagelos presentan nueve dobletes de microtúbulos con un par de microtúbulos centrales; mientras los centriolos y cuerpos basales tienen nueve tripletes de microtúbulos y ninguno en el centro. Además, cilios y flagelos están recubiertos por una extensión de la membrana plasmática, por lo que se consideran estructuras intracelulares. La hidrólisis del ATP, propiciada por la proteína dineína, permite el deslizamiento de los microtúbulos, lo que se traduce en el movimiento de cilios y flagelos.
Los cilios y flagelos son estructuras de locomoción en los eucariontes unicelulares; mientras que en los organismos pluricelulares desempeñan diferentes funciones; por ejemplo, los cilios que se encuentran en la superficie del epitelio respiratorio, ayudan a desplazar hacia el exterior, polvo, bacterias y células muertas que han quedado atrapadas en el mucus excretado por las células, y los que se encuentran recubriendo el interior de los oviductos de los mamíferos arrastran el embrión joven hacia la matriz.
El movimiento de moléculas y otros componentes celulares se realiza gracias a la presencia de actinas, que se encuentran a menudo asociadas con un gran grupo de proteínas llamadas miosinas, las cuales convierten la energía del ATP en movimiento. La función más relevante que desempeñan estas dos proteínas es la contracción muscular y una variedad de movimientos que incluye la división celular y los movimientos de las células sobre un sustrato o cuando una célula se desliza sobre otra; esto es especialmente importante en los procesos embrionarios que comprenden importantes migraciones de células.
Recientemente, se han descubierto secuencias de genes bacterianos que son semejantes a los que codifican para actina y tubulina en los eucariontes. Los filamentos de un sencillo citoesqueleto bacteriano controlan la longitud de la célula, su forma y el sitio donde se formará el tabique de división.
La presencia de hasta 50 diferentes proteínas es lo que caracteriza a los filamentos intermedios. Éstos se han agrupado en diversas clases, de acuerdo con su composición química y su localización. Las más abundantes son las queratinas que se usan para formar uñas, cuernos, pezuñas y pelo. En las células epiteliales hay abundancia de queratinas blandas asociadas con los desmosomas que mantienen la integridad de los epitelios.
Filamentos intermedios interesantes son los neurofilamentos, que se encuentran en las neuronas maduras, especialmente en el axón, y cuya función es el soporte de los axones, que en ocasiones alcanzan hasta un metro de longitud. Las láminas nucleares que se encuentran por debajo de la envoltura nuclear pertenecen también a este tipo de filamentos.