
![]()
![]()
Las células provienen de otras células, lo que biológicamente significa que no hay generación espontánea. ¿Cuáles son las diferentes formas en las que las células se dividen para dar lugar a células nuevas que garanticen la continuidad de las especies y el incremento en la diversidad? Esta sección trata sobre el ciclo de las células y los mecanismos celulares y moleculares que explican cómo se dividen. Un requisito indispensable para la división de la célula es que duplique su composición molecular y, en particular, su material genético. En procariontes se da por bipartición, mientras en eucariontes es por mitosis. La mitosis es parte del ciclo celular de las células eucariontes.
La reproducción no es un proceso continuo, sin embargo, la mayoría de las células se dividen a intervalos regulares y crecen en otros intervalos, constituyendo lo que se denomina un ciclo celular, es decir, una serie ordenada de acontecimientos que conducen a la replicación celular con una repetición alternante de periodos de crecimiento y división celular.
Este ciclo celular, al menos en eucariontes, se divide en cuatro fases secuenciales: una fase G1, en donde ocurren todas las funciones vegetativas como el crecimiento o aumento de la masa celular por duplicación de organelos y de macromoléculas y la producción de material celular o de secreción, entre otras; una fase S (síntesis), en donde se duplica el ADN, si la célula se prepara hacia una nueva división; una fase G2, en la que se terminan los preparativos para la reproducción, se sintetizan proteínas asociadas a los cromosomas y finalmente disminuye la actividad metabólica, y una fase M (mitosis o meiosis), en la que ocurre el reparto del material genético, seguida por la citocinesis o división del citoplasma.
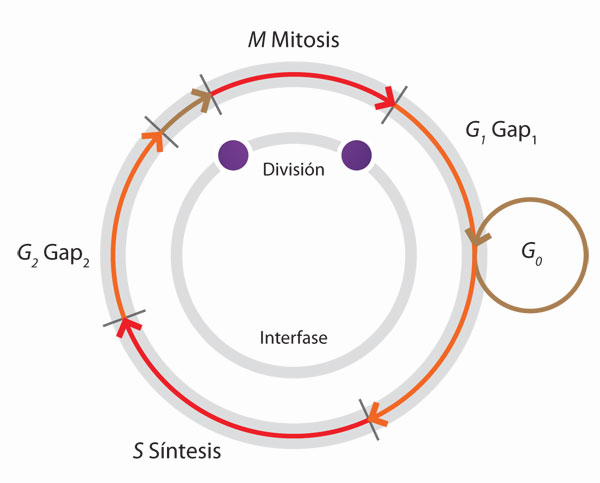
Las fases G1, S y G2 se incluyen en un conjunto llamado interfase. Las fases G1 y G2 permiten a la célula evaluar si las condiciones del medio externo e interno son las adecuadas para llevar a cabo la reproducción. No hay células que sigan este ciclo en forma continua ni una duración específica para cada una de estas fases; de hecho, la duración de la fase G1 puede variar en función de las condiciones internas y de las señales extracelulares, de forma tal que una célula en G1, al final de esta fase, entra en un punto que se conoce como de restricción o de inicio, durante el cual la célula evalúa su capacidad de completar el ciclo y "decide" si se duplica su ADN y sigue el proceso o si entra en una fase de reposo o G0, durante días, semanas, meses o años, antes de volver a reproducirse. Algunas células especializadas permanecen en esta fase en forma indefinida, otras en cambio, estimuladas por agentes mitógenos como hormonas, nutrientes o factores de crecimiento, pueden salir de G0, recomenzar el ciclo y dividirse.
Como todas las funciones de los sistemas vivos, el ciclo celular es también un proceso regulado. Las células eucariontes, en particular, han desarrollado un complejo sistema de control de este ciclo sobre la base de proteínas y el establecimiento de una serie de puntos de control (checkpoints) que aseguran que todos los componentes celulares estén presentes para que la célula pueda entrar en la siguiente fase; en estos sitios de control el ciclo es regulado por señales procedentes de otras células que pueden estimular o inhibir la proliferación celular, o bien verificar si el proceso previo se ha desarrollado de manera satisfactoria y, de no ser así, retrasar o impedir la progresión del proceso.
La regulación del ciclo celular es un proceso complejo que se efectúa como una cascada de reacciones de fosforilación y desfosforilación debidas a la acción de quinasas y fosfatasas, respectivamente. El agregar o quitar fosfatos a una enzima determina su cambio de conformación, lo que puede activar o inhibir su acción.
Las quinasas o cinasas que participan en la regulación del ciclo celular son enzimas que dependen de proteínas reguladoras denominadas ciclinas; cuando las ciclinas no están presentes, las quinasas no muestran actividad alguna. Las quinasas dependientes de ciclinas (CDK; de cyclin-dependent kinases), pueden asociarse con diferentes ciclinas y de esta manera se determina el tipo de proteína que se fosforila.
Existen tres tipos de CDK: las que participan en la fase G1, en la fase S y en la mitosis.
La duración del ciclo, el número de veces que puede repetirse y el número de células que pueden coexistir depende del tipo celular y se regula por condiciones internas y externas. El ritmo de la muerte celular también es un proceso regulado; así, en células que ya han realizado un determinado número de divisiones, presentan alguna anormalidad o cuya presencia y función ya no son necesarias, o ante la presencia de señales de proliferación anormales ocurre un fenómeno llamado apoptosis o muerte celular programada; ésta implica la destrucción de la célula mediante una cascada proteolítica que se origina al activarse proteínas de una familia de proteasas llamadas caspasas. Como resultado de su actividad, se destruyen diversas proteínas, muchas de ellas constituyentes de la membrana nuclear y se activan otras proteínas que finalmente degradan por completo los componentes celulares, para que sus residuos sean reaprovechados por las células vecinas o por macrófagos. Todo esto con el fin de mantener el número de células del organismo en cierto límite y, así, el buen funcionamiento del mismo.
No obstante, algunas células pueden escapar de estos sistemas de control y dividirse en forma incontrolada, amontonándose unas sobre otras hasta causar la muerte del organismo. Estas células son llamadas cancerosas, y se comportan de esta forma porque han perdido la capacidad de autodestruirse, quizás por una falla genética que bloquea los genes activadores del proceso apoptósico, como algunas proteínas de la familia Bcl-2, Bax, Bad o por producir proteínas como los llamados factores de supervivencia o telomerasas, que inhiben el proceso de muerte celular programada.
Mitosis | Este mecanismo de reproducción celular propio de células somáticas ocurre en la fase M del ciclo celular e incluye diferentes estadios de división nuclear y citoplásmica; en un relativamente corto tiempo, la célula que se reproduce debe repartir su contenido entre las dos células hijas a las que dará origen.
La mayor parte del tiempo, la mayoría de las células permanecen en lo que se conoce como interfase, que como ya hemos mencionado incluye las fases G1, G2 y S del ciclo celular. La aparente continuidad de forma e incluso de tamaño podría hacer confundir a la interfase con un periodo de reposo; sin embargo, es durante este lapso cuando se llevan a cabo todos los preparativos para la división celular, si ése es el caso de la célula, además de la realización de las funciones normales de la misma; así debe quedar muy claro que las células en interfase lo que menos hacen es reposar.
La consecuencia de la fase M es la distribución en forma precisa y equitativa del material genético de la célula que se reproduce y que ya ha sido replicado durante la fase S, de manera que al finalizar el proceso, las células hijas reciban una copia idéntica del genoma de la célula original.
Para que dé inicio la fase M, la célula deberá haber pasado por el punto de restricción y mediante la acción de proteínas como las CDK habrá desencadenado una cascada de fosforilaciones de proteínas que determinan el principio de la mitosis.
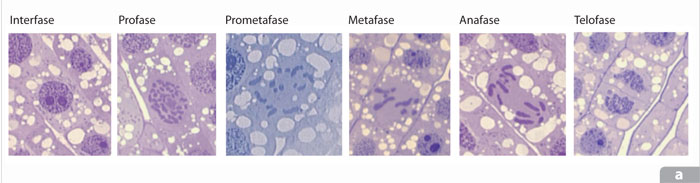
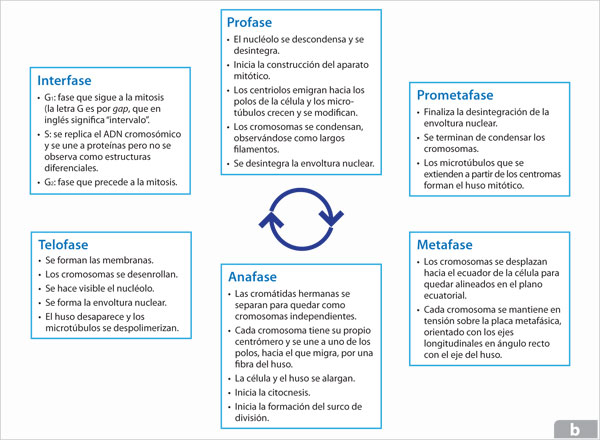
La primera señal visible del inicio de la mitosis es la progresiva compactación de las finas hebras de la cromatina replicada o cromátidas hermanas que permanecen unidas desde que se replica cada cromosoma, mediante la acción de cohesinas, unos complejos de proteínas de subunidades múltiples y de cuya actividad depende el éxito de la condensación y de la segregación posterior. El proceso de compactación continúa hasta la formación de los cromosomas mitóticos en cuya condensación completa participan condensinas, complejos de proteínas que al ser fosforiladas algunas de sus unidades por las M-Cdk desencadenan el ensamblaje de estas proteínas y el ADN y con ello la condensación del cromosoma.
En las células animales es necesario que durante la interfase se duplique el centrosoma, una estructura que contiene un par de centríolos y actúa como centro organizador de microtúbulos y que, al inicio de la mitosis, se separen los dos pares de centriolos para formar otro centro organizador de microtúbulos radiales llamado áster.
Una vez condensados los cromosomas se ensamblan en secuencia dos maquinarias del citoesqueleto, que realizarán los procesos de mitosis y citocinesis. Por una parte, se forma un aparato o huso mitótico bipolar conformado por los microtúbulos y algunas proteínas motoras que interactúan con ellos, a partir de cada uno de los centros organizadores, que entonces ya se han separado y empezado a migrar hacia polos opuestos del citoplasma. Esta serie de cambios que ocurren en la célula se incluyen en la primera fase de la mitosis: la profase.
La envoltura nuclear se rompe y los cromosomas son capturados por las fibras del huso, uniendo la región del centrómero de cada cromosoma con el cinetocoro de cada fibra, estableciendo así la unión de los microtúbulos con cada una de las cromátidas de los cromosomas. El aparato mitótico comienza realmente a ensamblarse. Esta fase se conoce como prometafase.
Al irse estableciendo la unión de los cromosomas con las fibras del huso es posible observar el alineamiento de los primeros formando una placa metafásica en el ecuador de la célula. Se trata de una etapa en la que los cromosomas oscilan suavemente, pues son arrastrados hacia delante y hacia atrás por las fibras de ambos lados del huso y esperan la señal para separarse, lo que ocurrirá cuando todos los cromosomas hayan alcanzado la adhesión bipolar, es decir, cuando los microtúbulos de ambos lados del huso hayan alcanzado los dos lados de la región centromérica del cromosoma y se hayan unido a las dos cromátidas. Estos acontecimientos caracterizan a la metafase.
La anafase da inicio con el rompimiento del enlace de la cohesina que mantiene unidas a las cromátidas hermanas. Este proceso se desencadena por la activación del complejo promotor de la anafase que inactiva a la ciclina M y detiene la actividad de la M-Cdk, e inactiva a la proteína inhibidora de la separasa, una proteasa que corta una subunidad del complejo de la cohesina permitiendo que se separen las dos cromátidas que comienzan a migrar hacia polos opuestos. Esta migración de cromátidas va acompañada de un acortamiento de los microtúbulos en el lugar de unión al cromosoma y por una despolimerización de los microtúbulos desde los dos polos del huso. El huso comienza a alargarse y los microtúbulos se deslizan unos sobre otros por efecto de algunas proteínas motoras.
Al final de la anafase, cuando los cromosomas hijos se han separado comienzan a descondensarse formando grupos iguales en cada polo de la célula; los fragmentos de envoltura nuclear se asocian con la superficie de los cromosomas y se fusionan para reconstruirla. En este proceso se incorporan los complejos de los poros nucleares y se restablece la continuidad de la envoltura nuclear con el retículo endoplásmico. Los poros nucleares bombean las proteínas nucleares al interior, el núcleo se expande, el nucléolo se reorganiza y los cromosomas terminan su descondensación. Con esta etapa, la telofase, la mitosis se completa y sólo resta la división citoplasmática o citocinesis.
La citocinesis en realidad comienza en la anafase, y requiere para su realización del ensamblaje de una segunda maquinaria del citoesqueleto, que se inicia con la aparición de un surco de segmentación en algunas células o un anillo contráctil en otras, constituidos de filamentos de actina, miosina II y otras proteínas estructurales que van ensamblándose por debajo de la membrana plasmática y que se van contrayendo para dividir a la célula por la mitad por una especie de estrangulamiento. Durante este proceso se adiciona membrana a partir de fusión de vesículas intracelulares para compensar el aumento de superficie celular que acompaña a la división. La formación del surco y sobre todo el plano en que debe formarse se determina por el eje del aparato mitótico y su ensamblaje y progresión dependen de la disminución de actividad de la M-CDK y de la activación de proteínas polares, aunque este último mecanismo no se conoce muy bien. En células vegetales la citocinesis ocurre por la formación de una placa celular a partir del fragmoplasto, una estructura formada por microtúbulos.
Durante la mitosis y la citocinesis, la célula también efectúa el reparto de los organelos, algunos de ellos o fragmentos de ellos se asocian con los microtúbulos del huso y son transportados hacia uno y otro polo, para luego reensamblarse.
Meiosis | Este tipo de división celular también corresponde a la fase M del ciclo celular, pero a diferencia de la mitosis es propio de células germinales, da origen a células hijas con un número cromosómico haploide y con un contenido genético diferente en cada una de ellas.
Considerando que los eventos citoplasmáticos que contempla una división celular en eucariontes, como la duplicación del centrosoma, el ensamblaje del huso, la ruptura de la membrana nuclear y su posterior reestructuración y la formación del surco de segmentación, por mencionar algunos, ocurren en forma similar en la meiosis, centraremos la atención sobre el comportamiento de los cromosomas, pues es allí donde se encuentran las mayores diferencias entre ambos tipos de división celular.
Tal como ocurre en la mitosis, antes de la meiosis, los cromosomas se replican durante la fase S y permanecen unidos a sus copias gemelas formando cromátidas hermanas.
Al inicio de división I de la meiosis, los cromosomas homólogos duplicados, versiones muy parecidas pero no idénticas de ADN, se reconocen y se aparean, tal vez mediante interacciones de pares de bases de ADN muy abundantes y ampliamente distribuidas a lo largo de cada cromosoma. Cuando este apareamiento ocurre se forma una estructura denominada bivalente que tiene cuatro cromátidas unidas físicamente en puntos de posición y número variable llamados quiasmas, lo que permite la recombinación genética, un proceso en el cual un fragmento de la cromátida materna puede intercambiarse por el fragmento correspondiente en la cromátida paterna. Este apareamiento puede durar horas, días o años y corresponde a una larga profase I. Esta fase se divide en cinco etapas: leptoteno o de condensación de pares de homólogos duplicados; zigoteno o inicio del desarrollo del complejo sinaptonémico entre cromátidas hermanas de cada bivalente; paquiteno o ensamblaje de las sinapsis y diploteno o de terminación de las sinapsis cuando es posible observar los quiasmas. Este proceso de recombinación está catalizado por nódulos de recombinación, constituidos por complejos proteicos que marcan, cortan y pegan diferentes zonas de los cromosomas homólogos.
Los cromosomas bivalentes liberados al disolverse la envoltura nuclear se asocian con los microtúbulos y al final de la metafase I se alinean en el ecuador del huso mitótico, para separarse y comenzar a migrar hacia polos opuestos marcados por los organizadores del huso durante la anafase I para, finalmente, reconstituir dos células en la telofase I y la citocinesis. El resultado de esta primera división meiótica origina células haploides en cuanto a dotación de ADN, pero no en cuanto al número de cromosomas, pues como las dos cromátidas hermanas se comportan como una unidad, cada célula hija en esta división se ha quedado con dos copias de alguno de los homólogos; sin embargo, estas copias ya no son idénticas como resultado de la recombinación. Se requiere de otra división celular, pero esta vez no irá mediada por una fase S, es decir, no habrá duplicación de ADN.
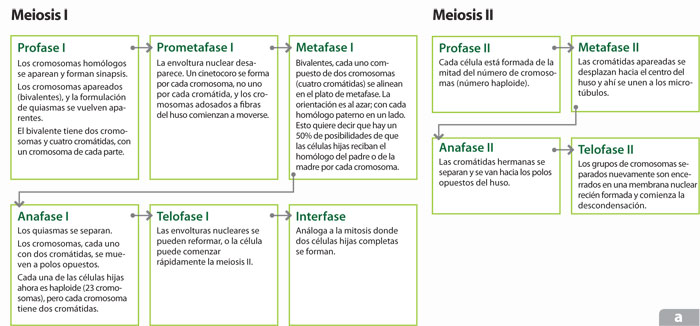
En la división II de la meiosis, nuevamente los cromosomas constituidos por cromátidas hermanas se asocian a las fibras de nuevo huso mitótico y se alinean en el ecuador del mismo, habiendo pasado por una profase II para que, al final de la metafase II, comience la separación de las cromátidas hermanas en la anafase II mediante un proceso llamado disyunción, para dar lugar a una telofase II y una citocinesis II que origine células haploides en contenido cromosómico y dotación de ADN, los gametos. Dado que esta segunda división ocurre en las dos células resultantes de la primera división meiótica, el resultado final de la meiosis son cuatro células hijas.