
![]()
![]()
El planteamiento de que el material hereditario es una partícula abrió la posibilidad de su identificación química y molecular. Primero se descubrió el ADN y luego se determinó su papel en la herencia. El descubrimiento de la composición atómica y de su arreglo en el espacio se debió a una serie de de hallazgos: la composición de bases nitrogenadas; la disponibilidad de la metodología de análisis molecular por rayos X; los datos disponibles, tanto de genética como de biología celular, y los conocimientos químicos de la época, entre otros, llevaron a conocer en detalle la arquitectura molecular del gen.
A pesar de la evidencia del parecido de los hijos con sus padres desde hace miles de años, las causas de esa similitud pasaron inadvertidas durante mucho tiempo. Hipócrates (400 a.C.), por ejemplo, señalaba que en todas las partes del cuerpo se producían semillas (pangénesis), mismas que se colectaban y trasmitían a la progenie en el momento de la concepción, y que esas semillas eran la causa de que determinadas características fueran similares a las de los progenitores.
Las ideas de Hipócrates fueron aceptadas y rechazadas durante los 2 000 años siguientes, hasta que la invención del microscopio en el siglo XVII permitió la observación de los espermatozoides. Aunque la interpretación de los mismos fue en un principio errónea, se inició una nueva manera de estudiar los fenómenos de reproducción y de herencia biológica.
El microscopista holandés Antony van Leeuwenhoek, uno de los descubridores del espermatozoide en 1678, creía que el espermatozoide era un animal parásito que vivía en el semen (de ahí el nombre de spermatozoa, que significa animales del semen). Originalmente, Leeuwenhoek no encontró una relación entre el espermatozoide y la reproducción del organismo, hasta que después pretendió que dentro del espermatozoide se encontraba ya preformado un ser humano al cual se llamó homunculus (hombre pequeño). El papel de la madre era meramente pasivo, al proporcionar los elementos necesarios para su desarrollo dentro del útero; con ello se regresaba a los conceptos promulgados por Aristóteles 2 000 años atrás. Además de esta escuela que puede llamarse espermista, existió por la misma época la de los ovistas, para quienes el óvulo era el responsable único de las características humanas.
Los primeros estudios sistemáticos sobre la herencia de los caracteres fueron realizados entre 1761 y 1766 por Joseph Kölreuter, quien hizo una serie de cruzas con plantas de tabaco y encontró que la progenie era más o menos igual entre sí y que presentaba estados intermedios entre los de los progenitores. Esto lo llevó a concluir que ambos padres participaban por igual en la distribución de las características físicas de la progenie. Sus observaciones apoyaron la teoría de la mezcla de las características hereditarias. De acuerdo con esta teoría, en cada generación las características se mezclaban para formar nuevas combinaciones. Esta idea, junto con la pangénesis, permitió que hasta el siglo XIX se pensara que las características de los individuos podían modificarse en el curso de una o dos generaciones.
En 1886, Gregorio Mendel (uno de los representantes más notables de una pléyade de "hibridadores", cuyas actividades experimentales tuvieron una enorme influencia en la instauración de las nuevas concepciones de la herencia) publicó una serie de experimentos por los que en la actualidad se le conoce como el "padre de la genética".
Mendel trabajó entre 1856 y 1863 en el cruzamiento de 63 variedades del chícharo Pisum sativum, planta cuya flor tiene una forma que favorece la autofecundación, por lo cual los experimentos de cruza podían controlarse con facilidad. Realizó miles de cruzamientos entre las plantas de chícharo, los cuales registró minuciosamente, incluyendo datos cuantitativos susceptibles de ser analizados estadísticamente. En 1866 publicó su trabajo intitulado Versuche über Pflanzen hybriden (Experimentos sobre hibridación de plantas) en una revista poco conocida de la ciudad de Brno, por lo cual su trabajo permaneció sin ser reconocido hasta finales del siglo, cuando fue redescubierto de manera independiente por tres biólogos interesados en la genética de las plantas: Hugo de Vries, de Holanda; Carl Correns, de Alemania, y Erich von Tschermak, de Austria.
Cuando Mendel inició sus trabajos sobre la herencia en el chícharo, se desconocía la existencia de los cromosomas y el mecanismo de la meiosis; sin embargo, determinó que existían unidades discretas de herencia que él llamó bildungfähigen elemente (elementos constructores de formas, posteriormente denominados genes) y fue capaz de predecir su comportamiento durante la formación de los gametos. Con ello demostró que la herencia no era, como se pensaba, una mezcla de sangre o de algún otro tipo de sustancia con la cual contribuían ambos progenitores. Por el contrario, encontró que en cualquier característica que él estudiaba, existían para cada individuo dos partículas o elementos, uno de ellos heredado de la madre y el otro del padre. Cada uno conservaba su individualidad, de tal modo que al momento de producirse los gametos, éstos obtenían uno solo de los elementos.
Las primeras cruzas efectuadas por Mendel fueron monohíbridas, es decir, mediante el intercambio de polen de plantas pertenecientes a líneas puras, que sólo variaban en una característica contrastante: semillas lisas-semillas rugosas; tallos largos-tallos cortos; semillas amarillas-semillas verdes, etc. Se denomina línea pura a aquella que conserva la misma característica a través de varias generaciones formadas por autofecundación.
La generación original o generación parental (P) se cruzaba para dar origen a la primera generación filial (F1); ésta se dejaba autofecundar para que se produjera la segunda generación filial (F2).
La cruza de una generación parental constituida por un individuo poseedor de tallos largos y uno de tallos cortos daba como resultado una filial 1 formada por plantas que sólo tenían tallos largos. Cuando Mendel dejó que estas plantas se autofecundaran obtuvo la filial 2, que presentó plantas con tallos largos y plantas con tallos cortos en una proporción aproximada de 3:1.
Mendel hizo igualmente experimentos de cruza con las plantas que tenían las otras seis características contrastantes, y en todos los casos obtuvo los mismos resultados: la F1 siempre mostraba sólo una de las dos características, mientras la F2 mostraba las dos en una proporción de 3:1.
Los experimentos de cruza que realizaba para obtener la filial 1 los hacía polinizando la planta de tallo largo con polen de planta de tallo corto o viceversa, procedimiento que se denomina cruza recíproca, y los resultados que obtenía eran siempre los mismos, demostrando con esto que no dependían del sexo.
De las cruzas monohíbridas de Mendel se derivan dos postulados y una ley:
• Cada carácter está controlado por un par de elementos que existen en el individuo.
• Dominancia/recesividad: si sólo una de las características aparece en la filial 1, entonces ésta debe ser dominante sobre la otra que se denomina recesiva.
• Ley de la segregación. Durante la formación de los gametos, cada elemento se separa o segrega al azar, de tal manera que cada uno de ellos tiene la misma probabilidad de recibir uno u otro.
Lo anterior explica perfectamente lo observado en las cruzas monohíbridas de Mendel. La planta con tallo largo posee dos elementos iguales AA, mientras que la de tallo corto tiene aa. Los gametos de ambas plantas recibirán sólo un elemento A o a. Al unirse, la filial 1 tendrá Aa. La mitad de los gametos de la F1 llevará el factor A y la otra mitad portará a.
Al mezclarse los gametos femeninos A y a con los gametos masculinos A y a, se obtienen las siguientes combinaciones: AA, Aa y aa. Si aceptamos que el carácter de tallo largo es dominante sobre el tallo corto, entonces la filial 1 será toda de tallo largo, mientras la filial 2 será de tallo largo y tallo corto, y mostrará una proporción de 3:1.
Mendel también hizo experimentos en los cuales demostró el comportamiento
de dos características. Estos cruzamientos dihíbridos fueron la base para la formulación de su segunda ley: la segregación o separación independiente de los elementos. Si se cruza una planta con semillas lisas de color amarillo, con una que produzca semillas rugosas de color verde (ambas pertenecientes a una línea pura), se obtendrá una F1 formada únicamente por plantas con semillas lisas y amarillas, demostrando con ello que las características lisa y amarillas son dominantes.
Si a la F1 se le permite autofecundarse, la F2 mostrará aproximadamente las siguientes proporciones: 9/16 de las plantas F2 expresarán los caracteres liso y amarillo; 3/16, rugoso y amarillo; 3/16, liso y verde, y 1/16, rugoso y verde.
Para entender estos resultados es necesario pensar en la cruza dihíbrida como si ocurrieran dos cruzas monohíbridas independientes. En cada cruza monohíbrida los resultados de la F2 son: 3/4 de plantas con el carácter dominante liso y 1/4 de plantas con el carácter recesivo rugoso; de igual manera, se obtienen 3/4 de semillas amarillas y 1/4 de semillas verdes; si multiplicamos ambos resultados tendremos:

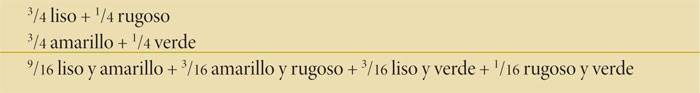
De acuerdo con estas observaciones, Mendel emitió su segunda ley que dice: durante la formación de los gametos, cada par de elementos se segrega independientemente del otro.
Si consideramos que la generación parental es AABB para las plantas con semillas lisas y amarillas y aabb para los progenitores con semillas rugosas y verdes, los gametos correspondientes a ambos padres serán AB y ab, respectivamente. Cuando los gametos se unen, se obtiene la F1 cuya fórmula es AaBb.
Cada progenitor de la F1, de acuerdo con la segunda ley, tendrá los gametos siguientes: AB, Ab, aB y ab.
¿Por qué tenemos estos resultados? Si consideramos cada par en forma independiente, podemos formular la siguiente pregunta: ¿qué probabilidad hay de que el gameto tenga A? La respuesta será 50%. ¿Qué probabilidad existe de que el gameto lleve B? La respuesta será la misma: 50%. Se procede de la misma manera con el par Bb. Si multiplicamos los resultados obtenidos estaremos en condiciones de predecir el tipo de gametos que tendrá cada progenitor de la F1:

Si la F1 se autofecunda, cada uno de los gametos de los progenitores tienen la misma probabilidad de unirse con otro; los resultados pueden demostrarse de manera gráfica con un cuadro de Punnet, llamado así en honor de Reginald Punnet, quien utilizó este método para demostrar los resultados de la cruza monohíbrida.
En la actualidad usamos diversos términos que favorecen la comprensión de los conceptos básicos de genética. Características tales como el color de una flor o la formación de una proteína cualquiera son expresiones de la información contenida en los genes. A la expresión física de los genes se le denomina fenotipo de un individuo. El fenotipo está determinado por los alelos, que son las formas alternativas de un gen. Por ejemplo, el tamaño de una planta de chícharo está determinada por el par de alelos A y a. Por conveniencia se acostumbra usar una letra mayúscula para representar el alelo dominante, mientras la minúscula representa el alelo recesivo.
El conjunto de alelos se denomina genotipo. Este término indica la composición genética de un individuo haploide o diploide, o también la de los gametos. Cuando ambos alelos son iguales, el individuo es homócigo u homocigoto, pero si los alelos son diferentes, se habla de un heterócigo o heterocigoto para ese par de alelos.
Los experimentos de Mendel revelaron que los organismos poseían un material que se trasmitía durante la reproducción y que regía el comportamiento de los rasgos que se heredaban.
Esta sustancia debía llenar los requerimientos de información necesaria para construir un organismo; replicación y trasmisión, para que el material genético se duplicara y pasara de los padres a la progenie; y variación por mutación que explicara la variabilidad entre los organismos.
Aunque los experimentos hechos por Mendel y otros genetistas a principios del siglo XIX eran consistentes con los postulados anteriores, el reconocimiento de la naturaleza química de la sustancia de la herencia necesitó de un gran trabajo de investigación que llevó poco menos de 100 años.
En 1868 Johann Friedrich Miescher, médico suizo, viajó a Alemania para trabajar bajo la dirección de Ernst Felix Hoppe-Sayler. Miescher desarrolló un método para aislar los núcleos de los leucocitos que formaban el pus. A pesar de las pequeñas cantidades de sustancia nuclear que obtenía, pudo establecer que era un material ligeramente ácido y con un alto contenido en fósforo. Llamó a esta sustancia nucleína.
En 1887, conforme los trabajos de investigación prosiguieron, se llegó a la conclusión de que las bases de la herencia radicaban en el núcleo. En 1905 Aaron Levene, del Rockefeller Institute de Nueva York, encontró que el ADN contenía cantidades iguales de moléculas denominadas nucleótidos, constituidas por un fosfato, un azúcar y una base nitrogenada; sin embargo, consideraba erróneamente que los grupos de nucleótidos se repetían una y otra vez, lo que dio origen a la hipótesis del tetranucleótido, para explicar la estructura del ADN. Se demostró, hasta ese momento, que la cromatina estaba constituida por proteínas y el ADN por cuatro nucleótidos; pero la escasa variación mostrada por el ADN lo hacía un candidato muy pobre para llevar la información genética.
En los medios científicos de la primera mitad del siglo XX privaba la idea de que las proteínas eran el material genético, pues estaban constituidas por 20 aminoácidos, lo que las hacía suficientemente variadas como para constituir la fuente de la información genética.
Fue en 1949 y 1953 cuando Edwin Chargaff y sus colegas, usando técnicas cromatográficas para separar los diferentes nucleótidos del ADN de distintos organismos y mediante métodos cuantitativos, determinaron las cantidades de las bases nitrogenadas que se encontraban en las diferentes muestras. Con ello se demostró que la hipótesis de Levene era incorrecta, al señalar que en los diferentes organismos se encontraba una cantidad variable de nucleótidos y que la cantidad de adenina era similar a la de la timina, y que la guanina igualaba en cantidad a la citosina, observación que más tarde se conoció como regla de Chargaff.
La primera pista que permitió el reconocimiento del ADN como material genético vino del descubrimiento del fenómeno de transformación hecho en 1928 por el médico inglés Frederick Griffith. Él estudiaba la bacteria Streptococcus pneumoniae, de la cual había aislado varias cepas diferentes; algunas de ellas eran cepas virulentas, que causaban neumonía en vertebrados, especialmente en humanos y ratones. Estas bacterias poseen una cápsula de polisacáridos bien desarrollada que hace que sus colonias tengan una apariencia lisa; las colonias de cepas no virulentas (no causan enfermedad ni tienen cápsula) presentan aspecto rugoso, lo que las hace vulnerables a los ataques de las células fagocíticas del sistema inmune de los animales.
Griffith observó que cuando se inyectaba la cepa sin cápsula S (de smooth, liso) a los ratones, éstos desarrollaban neumonía y morían; en la autopsia se encontraba siempre la cepa S en gran cantidad. Cuando se inyectaba la cepa R (de rough, rugoso) los ratones no mostraban signos de la enfermedad. Sin embargo, cuando se inyectó a los ratones la cepa S muerta por calor junto con la cepa R viva, los resultados fueron sorprendentes: los ratones murieron, a pesar de que la cepa virulenta estaba muerta. La sangre de los ratones muertos mostró una gran cantidad de bacterias con cápsula. La interpretación de los datos demostraba que algo de las bacterias virulentas muertas por calor había pasado a las no capsuladas y las había transformado en bacterias capsuladas virulentas, que conservaron esta característica a través de varias generaciones. Este proceso fue denominado "transformación" y la sustancia que lo había producido se conoció como principio transformante.
Las observaciones de Griffith permitieron que el equipo de Oswald Avery, Colin MacLeod y Maclyn McCarty, después de 10 años de investigaciones descubrieran la naturaleza química del principio de transformación.
Avery y su equipo trabajaron con las cepas R y S de P. Pneumoniae, con la intención de demostrar que el principio transformante tenía una estructura que lo acercaba más al ADN que a las proteínas. Para ello usaron sobre el principio transformante aislado enzimas que separadamente digerían ARN, ADN y proteínas. Cuando se usaban enzimas digestivas como la tripsina que degrada proteínas, la sustancia transformante no sufría cambio alguno (las bacterias R tratadas con ella se transformaban en bacterias S), descartándose de esta manera su naturaleza proteica. De la misma manera, la ribonucleasa que digiere el ARN tampoco mostraba efecto alguno sobre el principio transformante; pero cuando se usaban enzimas capaces de digerir el ADN, el principio transformante eliminaba su actividad biológica (las bacterias R no se transformaban en bacterias S) demostrándose con ello que el ADN era la sustancia que provocaba la transformación. La interpretación de los datos obtenidos por Avery y su equipo demostró que el principio transformante de Griffith y por lo tanto el material genético era, sin lugar a duda, el ADN. Los resultados de los experimentos fueron publicados en 1944.
Los experimentos hechos en 1952 por esta pareja de científicos proporcionaron una evidencia adicional de que el ADN era efectivamente el material genético. Sus trabajos se centraron en el bacteriófago T2 que infecta a la bacteria Escherichia coli. En la actualidad se sabe que el bacteriófago inyecta en la bacteria su ADN, el cual se replica y dirige la síntesis de proteínas virales a expensas de la maquinaria enzimática bacteriana.
En esa época se ignoraba cómo se reproducían los fagos, pero se sabía que estaban constituidos por proteínas y ácido nucléico. Sin embargo, se desconocía a cuál de las dos sustancias correspondía al material genético.
Alfred Hershey y Martha Chase diseñaron experimentos para resolver este problema. El uso de radioisótopos P32 y S35, que pueden incorporarse sólo al ADN y a las proteínas, respectivamente, les permitió comprobar en forma definitiva que el ADN era la sustancia inyectada por el virus y la que proporcionaba la información para sintetizar más moléculas de ADN y proteínas virales.
El descubrimiento de la estructura de la molécula de ADN es uno de los logros científicos más espectaculares de los últimos tiempos. Si bien se sabía desde hacía tiempo que el ADN estaba constituido por nucleótidos, se ignoraba la forma en que éstos se encontraban unidos para formar la estructura del ácido nucléico.
Los resultados obtenidos por Chargaff y una serie de análisis de difracción por rayos X, aplicados al ADN, primero por Linus Pauling y otros químicos, y después por Rosalind Franklin, quien trabajaba en el laboratorio de Maurice Wilkins, abrieron el camino para que en 1953, James Watson y Francis Crick propusieran el modelo de doble hélice para la molécula de ADN, que en principio no fue bien aceptada por los bioquímicos de Cambridge, debido a que se había obtenido sin hacer experimentos.
El modelo propuesto primero por Pauling y luego por Watson y Crick se refería a una estructura helicoidal formada por tres cadenas. Finalmente, Watson y Crick llegaron a la conclusión de que el ADN era una hélice formada por dos cadenas.
Las características fundamentales del modelo propuesto por Watson y Crick fueron las siguientes:
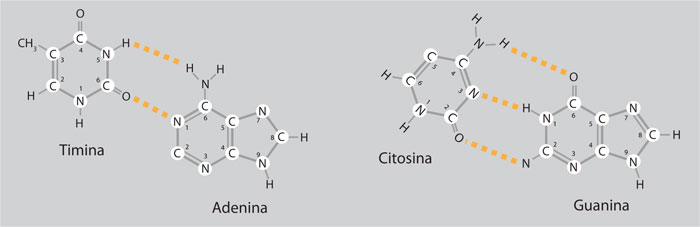
• Se trataba de una molécula formada por dos hebras enrolladas alrededor de un eje central, semejante a una escalera de caracol. A diferencia de un modelo previo en el cual las bases se encontraban hacia el exterior de la hélice, en el nuevo modelo los pasamanos de la escalera estaban constituidos por los grupos fosfatos y azúcares de tipo desoxirribosa y los peldaños de la escalera, por pares de bases; una purina se apareaba siempre con una pirimidina, es decir, la adenina con la timina y la guanina con la citosina.
• La estructura de la doble hélice se estabilizaba gracias a los puentes de hidrógeno que unían a las bases: dos para la pareja timina-adenina y tres para guanina-citosina. La regla Chargaff AT/CG, implicaba que ambas hebras de la doble hélice se complementaban, por lo que, si se conocía una de las hebras, era posible deducir la secuencia de la otra.
• Por el esquema de emparejamiento de bases, cada una de las hebras de la doble hélice giraba en dirección opuesta, generando antiparalelismo y polaridad a la molécula. La polaridad ocurre por el carbono del azúcar en el extremo de la cadena.
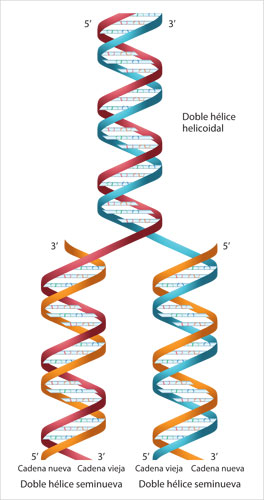
•La estructura de la molécula sugería la forma en la que podía duplicarse; de hecho, poco tiempo después de haberse publicado en 1953 el trabajo de Watson y Crick, los autores dieron a conocer la propuesta del modelo de la replicación semiconservativa. En este segundo artículo se señalaba además que en la secuencia de las bases nitrogenadas se encontraba la información genética, y que los cambios genéticos se debían a una modificación en la secuencia de las bases.

Después del descubrimiento de la estructura del ADN se desconocía el papel que desempeñaba el ARN, aunque se pensaba que podía ser un intermediario entre el ADN y las proteínas. Fue el físico ruso George Gamow quien se interesó en la relación entre el ADN y las proteínas. Gamow supuso que tripletes de bases servían para especificar ciertos aminoácidos.

En la década de 1950 Paul Zamecnik, quien laboraba en el Hospital General de Massachusetts en Boston, trabajando con hígado de rata, hizo el importante hallazgo de que las proteínas se sintetizaban en los ribosomas; posteriormente, junto con su colega Mahlon Hoagland, descubrió que los aminoácidos, antes de unirse en los ribosomas, estaban unidos a pequeñas moléculas de ARN. Esto concordaba con la hipótesis de Crick de que debía existir un "adaptador" (llamado ARN de transferencia) que llevaría los aminoácidos al sitio donde se sintetizaban las proteínas. Para cada aminoácido debía existir una molécula adaptadora.
Posteriormente, en 1960, se descubrió el ARN mensajero, que permitió la elaboración del modelo de síntesis de proteínas: el ARN mensajero con la información procedente del ADN llegaba a los ribosomas donde era "decodificado". Los ARN de transferencia, cada uno con su aminoácido específico, se unían al ARN mensajero, lo que aseguraba que el péptido tuviera un orden adecuado antes de formar los enlaces peptídicos.
Sin embargo, aún faltaba determinar las reglas del código que permitiera la traducción de una secuencia de ARN en una secuencia de aminoácidos. La pregunta básica era cuál de los 20 aminoácidos se incorporaría a la cadena peptídica, si sólo existen cuatro bases en el ADN. Para 20 aminoácidos harían falta por lo menos tres bases, aunque con éstas, la cantidad de combinaciones sería de 64, lo que implicaría que un aminoácido podría estar representado por más de un triplete.
Finalmente, Crick y Sydney Brenner demostraron que los tripletes eran la base del código genético. Mediante experimentos con sustancias mutágenas, pudieron insertar o eliminar bases de una secuencia de ADN, lo que provocaba en la proteína un cambio de estructura. Pero las inserciones y deleciones no tenían el mismo efecto si las bases eliminadas o insertadas eran una o dos, lo cual originaba un cambio que alteraba el orden de todos los aminoácidos del péptido. Sin embargo, tres deleciones o inserciones no producían las mismas perturbaciones en el orden de los aminoácidos; solamente eliminaban o agregaban un aminoácido a la cadena de péptido. Esto hizo que Crick emitiera la hipótesis de que el código genético debía ser un código de tripletes.
En 1961, la bioquímica francesa Marianne Grunberg-Manago descubrió una enzima que podía producir segmentos de ARN como AAAAAAAA... o GGGGGGG... Marshall Nirenberg y su colega Heinrich Matthaei utilizaron este hallazgo para sintetizar un ARN formado únicamente por uracio o poli-U que, al ponerse en contacto con ribosomas in vitro, empezó a formar una cadena compuesta sólo por el aminoácido fenilalanina. Se había descubierto que el triplete del código genético que codificaba para la fenilalanina tenía que ser UUU.
En los años siguientes se hicieron numerosos experimentos para descifrar los otros 63 tripletes. Muchos de los experimentos fueron hechos por Gobind Khorana, de la Universidad de Wisconsin. Para 1966 se estableció finalmente el papel que jugaban cada uno de los 64 tripletes del código genético. Nirenberg y Khorana recibieron en 1968 el Premio Nobel de Fisiología y Medicina.