Las actividades que llevan a cabo los organismos siempre tienen un efecto en su entorno. Por ejemplo, la transpiración de las plantas contribuye a aumentar la humedad en el ambiente, el ramoneo efectuado por un rebaño de cabras le da una apariencia al paisaje que seguramente sería diferente sin su presencia, pues la actividad constante de estos animales impide que se desarrollen normalmente las plantas, lo que propicia la desertización.
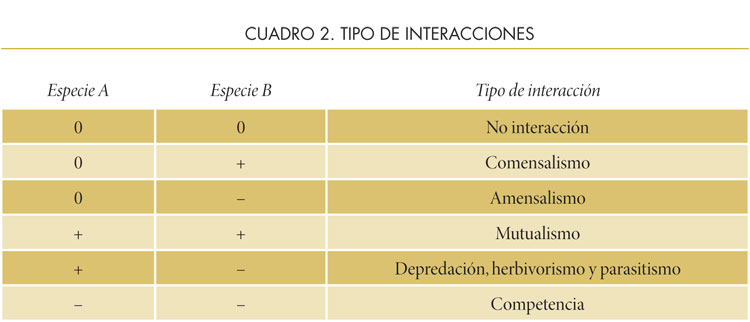
La interacción de los seres vivos no se da únicamente con el ambiente inanimado, sino también con organismos de otras especies. En una comunidad, existen diferentes tipos de interacciones, cuyos efectos pueden ser positivos (+), negativos (–) o neutros (0) para una o para ambas especies. Los principales tipos de interacciones son la competencia, la depredación (que incluye al herbivorismo, la depredación verdadera, y el parasitismo, el mutualismo y el amensalismo.
La competencia es un tipo de relación en que los organismos participantes resultan perjudicados, lo que se manifiesta en una disminución de la supervivencia, de su fecundidad o de su crecimiento. Se presenta en dos modalidades: intraespecífica e interespecífica. La primera ocurre entre los organismos de la misma especie, ya que utilizan los mismos recursos limitados, tales como alimento, territorio o pareja. La competencia interespecífica se da entre especies diferentes que ocupan nichos ecológicos semejantes y generalmente este tipo de competencia es menos intensa que la intraespecífica, debido a que en ésta los organismos tienen prácticamente los mismos requerimientos para su existencia.
La competencia interespecífica, de manera semejante a la intraespecífica, puede ocurrir de dos maneras: que los individuos de una especie obstaculicen directamente la supervivencia de los individuos de otra especie (competencia por interferencia), o bien, que una de ellas utilice los recursos de una manera más acelerada y eficiente que la otra (competencia por explotación), por lo que indirectamente impide el adecuado desempeño de esta última, a la que incluso puede provocar su extinción de la comunidad. Un ejemplo del primer tipo de competencia es la que existe entre dos especies de percebes (balanos): Chthamalus stellatus y Balanus balanoides, que viven adheridos a las rocas en ciertas regiones costeras. Aunque generalmente los adultos de estas dos especies se distribuyen en las rocas de manera diferenciada, existen zonas en las que coinciden los individuos jóvenes de Chthamalus con los adultos de Balanus; se ha observado que en la zona de confluencia de las dos especies, los adultos de Balanus asfixian o aplastan a los individuos jóvenes de Chthamalus.
La competencia por explotación ha sido comprobada experimentalmente en cultivos de algas microscópicas llamadas diatomeas. Éstas requieren de silicatos para construir sus paredes celulares. Cuando se cultivan dos especies juntas se observa que una de ellas utiliza con mayor eficiencia el silicato, reduciendo la disponibilidad de esta sustancia para la otra especie de diatomea, que al cabo de poco tiempo llega a desaparecer del cultivo.
Como se dijo antes, la competencia interespecífica se presenta cuando las especies se encuentran en condiciones ambientales similares; es decir, tienen nichos parecidos, pues sus requerimientos de recursos son muy similares. En ocasiones, el resultado de esta competencia es la exclusión competitiva o el desplazamiento de algunas especies de la comunidad llevado hasta el extremo de su desaparición local, causada directa o indirectamente por otra especie. No obstante, en ausencia de la especie "dominante", la que normalmente es desplazada tiende a ocupar los recursos que ocupaba la otra.
La exclusión competitiva tiene la categoría de principio, y fue establecido en 1934 por el ecólogo ruso Georgii F. Gause, quien hizo diversos experimentos con paramecios (protistas que viven en aguas con alto contenido de bacterias). Gause utilizó tres especies de Paramecium (P. bursaria, P. caudatum y P. aurelia), a los que cultivó primero de manera independiente, observando un crecimiento poblacional en forma de "S", debido a que se había llegado al límite en la capacidad de carga del ambiente. Posteriormente, cultivó de manera combinada a P. caudatum y P. aurelia, y encontró que P. aurelia causó la eliminación de P. caudatum del medio de cultivo por exclusión competitiva. Cuando colocó en el mismo medio de cultivo a P. bursaria y P. caudatum, advirtió que las dos especies coexistieron. Al hacer una observación más detallada, Gause se dio cuenta de que una de las especies se alimentaba de las bacterias que se encontraban en la superficie del cultivo, mientras la otra lo hacía de las levaduras del fondo; es decir, ambas se repartían los recursos, de tal manera que se evitó la competencia. A partir de estos descubrimientos, el investigador postuló otro principio: dos especies pueden coexistir, sólo si sus nichos son lo suficientemente diferentes; de lo contrario, una de las especies excluirá a la otra.
El principio de la diferenciación de nichos se ha confirmado tanto en situaciones experimentales como en observaciones de campo. Un ejemplo lo constituye el estudio que hizo Robert MacArthur sobre la distribución de cinco especies de aves del género Parus (herrerillo común) que habitan en bosques de Nueva Inglaterra. Él observó que aunque estas especies muestran muchas semejanzas morfológicas y de hábitat, presentan algunas diferencias en cuanto al tamaño y forma de sus picos, así como en sus preferencias alimentarias y de hábitat. Si bien las cinco anidan en la misma especie de árbol, mostraron ligeras desigualdades en cuanto al tipo de semillas y de insectos de los que se alimentaban, así como la zona de los árboles en donde anidaban. Es evidente que, a pesar de sus semejanzas, estas especies utilizan los recursos de una manera levemente diferente, por lo que prácticamente no compiten entre sí, ya que se dio una diferenciación de nichos.
A pesar de que el principio de exclusión competitiva está apoyado por muchas observaciones y comprobado en diversas situaciones experimentales, no puede considerarse como un principio que se aplique a todas las comunidades, ya que la dinámica de cada una dependerá del tipo de comunidad de que se trate y de las especies que la conformen. Existen comunidades en las que las especies con nichos similares no se traslapan, pues tienden a utilizar los recursos de manera desigual, por lo que no compiten de manera intensa.
Es necesario mencionar que actualmente existe discrepancia entre los ecólogos acerca de la importancia de la competencia en la determinación de la estructura de las comunidades, así como de las especies que las conforman. Puesto que la estructura actual de las comunidades es el resultado de un proceso evolutivo, es muy útil tomar en cuenta la competencia interespecífica como una de las fuerzas que modelaron las características actuales de dichas comunidades. Sin embargo, debe ser valorada conjuntamente con otro tipo de interacciones como la depredación y el mutualismo.
Se trata de un tipo de relación en la que uno de los integrantes se alimenta de otro organismo vivo, por lo que una de las especies es beneficiada (el depredador), mientras la otra resulta perjudicada (la presa). Las diferentes modalidades que presenta esta interacción incluyen la depredación verdadera, el herbivorismo, el parasitismo y los parasitoides.
En ésta, el depredador consume a las presas causando su muerte casi de manera inmediata. Dentro de este grupo se encuentran los grandes carnívoros, como jaguares, pumas, coyotes, lobos y leones; muchas especies de aves como águilas, halcones y búhos; tiburones blancos, orcas y delfines, etcétera. También se incluyen invertebrados como arañas, insectos adultos como los mántidos y larvas de diversos insectos. Las presas generalmente son organismos herbívoros como cebras, gacelas, conejos, ratones, muchas especies de insectos, etcétera.
Los depredadores consumen muchas presas durante su vida, por lo que afectan los parámetros poblacionales y, por tanto, el tamaño de sus poblaciones; pero al mismo tiempo, la abundancia de presas incide en las poblaciones de los depredadores. Lo anterior es particularmente notorio en los organismos que se alimentan de una sola especie. Como consecuencia, el tamaño de las poblaciones de los depredadores y las presas se regula mutuamente de manera cíclica a través del tiempo. Se han hecho registros en los que se determinó la fluctuación de las poblaciones de linces (Lynx canadensis) y las liebres (Lepus americanus). La abundancia de liebres determina el número de linces que pueden sobrevivir, pues si existe poca disponibilidad de alimento, muchos morirán. Al disminuir la cantidad de depredadores, la población de liebres aumenta su tamaño; por lo tanto, al haber mayor disponibilidad de presas, la población de linces empezará a recuperarse. Ambas especies ejercen mutuamente presiones selectivas muy intensas, por lo que a través del tiempo han desarrollado estrategias que permiten, a unos ser cazadores más eficientes, mientras a otros los ayuda para intentar el escape ante sus enemigos naturales.
De esta manera, unos depredadores han adquirido agilidad y velocidad para capturar eficientemente a sus presas, mientras otros prefieren pasar desapercibidos al imitar al entorno (camuflaje) o a otro organismo (mimetismo). El mimetismo es frecuentemente utilizado por depredadores agresivos que sorprenden fácilmente a sus presas al asemejarse a especies inofensivas. El mimetismo y el camuflaje también son recursos utilizados por ciertas especies, como la mariposa virrey (Limenitis archippus), que presenta un patrón de coloración muy semejante a la mariposa monarca (Danaus plexippus) cuyo sabor es desagradable, por lo que las aves evitan comerlas. Por otro lado, muchas especies venenosas han desarrollado colores brillantes y llamativos que sirven de advertencia a sus enemigos, lo que reduce la probabilidad de ser atacadas.
Consiste en el consumo de tejido vegetal vivo, y los organismos que se alimentan de ello se denominan herbívoros. Los herbívoros abarcan una gran variedad de seres vivos, entre ellos muchos insectos y vertebrados. Generalmente los herbívoros no se alimentan del 100% de los tejidos de la planta, sino que tienen preferencias por ciertas estructuras: hojas, tallos, semillas, flores, frutos o raíces. Existen herbívoros que se alimentan de un número variado de especies vegetales, por lo que se les denomina generalistas, mientras que los especialistas interaccionan con una especie en particular, como las orugas de la mariposa Battus philenor que se alimentan sólo de la planta Aristolochia reticulata.
En la mayoría de los casos, la acción de los herbívoros tiene efectos negativos en las plantas, ya que al comer parte de sus tejidos pueden disminuir su velocidad de crecimiento o bien provocar una reducción de su éxito reproductivo. No obstante, los efectos del herbivorismo dependerán de varios factores, entre ellos la edad de la planta y la época del año en la que ocurre la depredación: los efectos del forrajeo por parte del ganado en los pastos de una pradera serán más intensos en la temporada de secas que de lluvias, pues en este periodo los pastos se recuperan más rápidamente. No obstante, en el contexto global de una comunidad, el herbivorismo moderado aparentemente puede incrementar la diversidad, al favorecer la dispersión de semillas lejos de la planta parental que está siendo atacada, lo que permite el establecimiento de plántulas de otras especies; o bien, al reducir el vigor de especies vegetales dominantes, se da lugar a la coexistencia de plantas con menor habilidad competitiva. Así, los herbívoros pueden modificar la diversidad de la comunidad.
Es necesario resaltar que esta interacción produce presiones selectivas mutuas en las especies involucradas, lo que ha favorecido su evolución conjunta (coevolución). Por su parte, las plantas han desarrollado mecanismos de defensa contra el ataque de los herbívoros, que van desde los mecánicos, como la producción de espinas, pubescencias y epidermis esclerotizada; los químicos, como la síntesis de resinas, y toxinas como los alcaloides y los glucósidos cardiacos, hasta los temporales, que tienen que ver con la producción de semillas y hojas nuevas, en periodos de tiempo diferentes al surgimiento de estados larvarios de los insectos que las atacan.
Como respuesta a estas defensas, los herbívoros también han desarrollado estrategias para evadirlas; cuando las plantas producen toxinas, algunos insectos evitan el consumo de las partes de la planta que las contienen. Por ejemplo, los áfidos (pulgones) Myzus persicae no comen las hojas de la planta de tabaco que produce nicotina, que es un alcaloide sumamente tóxico. Otras especies de herbívoros poseen un sistema excretor muy eficiente que evita la acumulación de compuestos tóxicos o presentan enzimas que los degradan y de esta manera no interfieren con el metabolismo del insecto.
Es un tipo de depredación donde el parásito, al igual que el herbívoro, se alimenta de su presa o huésped, causándole daño, pero sin matarla de manera inmediata. Generalmente, los parásitos se asocian con muy pocos huéspedes durante su vida, de ahí que se establezca una relación más estrecha entre ellos, que entre los depredadores verdaderos y sus presas.
El parasitismo se halla sumamente extendido en la naturaleza, pues prácticamente no existe un individuo que esté libre de al menos una especie de parásito. Debido a que aproximadamente la mitad de las especies en el planeta son parásitos, es evidente la importancia que tienen estos organismos en la regulación del funcionamiento de las poblaciones y en la evolución biológica. Desde el punto de vista humano, el parasitismo adquiere gran importancia, ya que es la causa de muchas de nuestras enfermedades y de las que padecen las especies de interés económico. Las especies parásitas incluyen a todos los virus, muchas bacterias, protistas, invertebrados —como algunos gusanos e insectos—, plantas como el muérdago, hongos —como el huitlacoche Ustilago— e incluso vertebrados, como algunas especies de aves y murciélagos hematófagos.
Existen varias clasificaciones de los parásitos. Una de ellas es la que distingue dos grupos: endoparásitos y ectoparásitos. Los primeros viven dentro de sus huéspedes: tenias, lombrices intestinales, amibas, Giardia, etcétera. Algunos de ellos, como los virus y algunas bacterias, son muy numerosos y se multiplican directamente en el interior de sus huéspedes, invadiendo sus células y afectando el metabolismo celular; mientras otros viven en las cavidades corporales, como las tenias y lombrices que habitan en el intestino de varios animales incluyendo el humano.
Los ectoparásitos viven en el exterior de su huésped, generalmente asociados con la piel, uñas, plumas, etcétera. Ejemplos de estos organismos son los ácaros, chinches, pulgas y piojos. Los piojos pasan todo su ciclo de vida en un huésped y la transmisión a otro suele darse por contacto físico directo entre ellos. Otros parásitos pasan los primeros estados de vida (huevo y larva) en un organismo, mientras que los adultos —mediante el salto— invaden a un nuevo huésped.
Los parásitos tienen ciclos de vida complejos, generalmente con una fase de dispersión que incluye un huésped intermediario, en el que pasan parte de su ciclo de vida y el cual actúa como vector para conducirlo al huésped final; en otros casos, los parásitos utilizan estructuras dispersoras, como esporas y quistes, que presentan cierta resistencia ante factores ambientales. Por ejemplo, algunos protistas como el Plasmodium, causante de la malaria o paludismo, para completar su ciclo de vida deben ser transferidos a su huésped final —humanos y otros mamíferos— a través de la picadura del mosquito Anopheles —huésped intermediario.
También muchas especies parasitan a las plantas: hongos como las royas y las cenicillas que atacan las hojas, y plantas como el muérdago que viven a expensas de los nutrientes de otras plantas. El caso más extremo de parasitismo por parte de plantas es el de Rafflesia arnoldii, especie cuyo cuerpo vegetativo se sitúa en el interior de la planta huésped, de la cual obtiene directamente todos sus recursos. Un dato curioso es que R. arnoldii produce la flor más grande que se conoce (hasta un metro de diámetro).
Es importante mencionar que la transmisión de los parásitos entre los individuos de una población se facilita en aquéllas con una alta densidad, como es el caso de las poblaciones humanas, en las que fácilmente se propagan enfermedades causadas por parásitos como el sarampión, el virus de inmunodeficiencia humana (VIH), que se propagó rápidamente desde su aparición, y más recientemente el virus de la gripe AH1N1.
Estos organismos generalmente afectan la supervivencia, el crecimiento y la fecundidad de los huéspedes. Por ejemplo, muchas enfermedades infecciosas en las sociedades humanas son causa importante de la mortalidad infantil, por lo que el parasitismo puede afectar la densidad de las poblaciones. Asimismo, los parásitos también pueden afectar la capacidad competitiva de las especies. Por ejemplo: la avena, cuando es parasitada por un nemátodo que afecta sus raíces, está en desventaja frente a la cebada. Por lo tanto, aunque sus efectos sean indirectos, los parásitos influyen en la dinámica de las comunidades.
Debido a que la interacción entre parásitos y huéspedes ejerce una presión selectiva mutua, es muy probable que estas especies coevolucionen. Un caso bien documentado es el del conejo europeo Oryctolagus cuniculus, introducido en Australia en la primera mitad del siglo XX, y el virus que causa la enfermedad llamada mixomatosis en la especie O. brasiliensis, pero de una manera no letal. Debido a que en Australia O. cuniculus se convirtió en plaga, en 1951 se introdujo el virus con el fin de controlar el tamaño de sus poblaciones. Al principio, el virus era letal para O. cuniculus, causando una gran mortalidad en su población (más del 90%). Al cabo de ocho años, las siguientes generaciones de virus fueron menos agresivas, y a su vez los conejos habían creado mecanismos de resistencia, por lo que ahora la mortalidad era menor al 30%. Este estudio respalda la afirmación de muchos ecólogos en el sentido de que cuando las especies evolucionan juntas, las relaciones parasitarias tienden a atenuarse. Es decir, la virulencia del parásito disminuye y los parasitados desarrollan mayor resistencia contra el parásito; de esa manera ambos aseguran su existencia.
Se refiere a un grupo de insectos a los que pertenecen ciertas avispas (himenópteros) y moscas (dípteros), cuyos adultos son de vida libre, pero sus larvas parasitan a otras especies de artrópodos. Este tipo de interacción es específica, pues los parasitoides atacan a determinados huéspedes. Las hembras buscan activamente a su huésped para oviponer en él, de tal modo que cuando emerjan las larvas se alimenten de su víctima. Debido a lo anterior, los parasitoides han sido empleados como un mecanismo de control biológico alternativo de ciertas plagas de insectos que afectan plantas de interés económico, evitando con esto el uso de insecticidas, que generalmente acarrean problemas de contaminación y toxicidad para el humano y otras especies.
Hasta aquí, se ha caracterizado de manera general cada una de las relaciones de depredación. Sin embargo, no se ha hecho referencia al papel de estas relaciones en el seno de las comunidades, en donde se dan interacciones complejas entre especies. Los resultados de los estudios encaminados a averiguar el papel de la depredación en la composición de las comunidades, indican que en algunas comunidades acuáticas la depredación ayuda a mantener una diversidad de especies mayor que si no hubiera depredadores.
Un caso bien estudiado es el de la depredación que ejerce la estrella de mar del género Pisaster, que se alimenta de mejillones. En presencia del depredador se mantienen poblaciones de mejillones de diferentes especies; mientras en su ausencia una especie de mejillón se desarrolla excesivamente, lo que inhibe el establecimiento de las otras poblaciones competidoras. La misma situación se ha observado en comunidades terrestres, en donde la presencia de un herbívoro mantiene una riqueza de especies de pastos mayor que cuando está ausente. Los anteriores son ejemplos de especies clave o angular, cuyas actividades ejercen un papel significativo en la determinación de la estructura y el funcionamiento de la comunidad. La analogía con la piedra angular o clave, se refiere a que si se remueve de un arco, éste se derrumba; de la misma forma, la remoción de una especie transforma a la comunidad.
No obstante se ha observado que si la depredación es intensa, el resultado será una disminución de la diversidad, pues las especies más vulnerables tienden a desaparecer, mientras sólo permanecerá aquella que soporte mejor la depredación. Por ejemplo, si una comunidad de pradera en la que se encuentran diferentes pastos es sobrepastoreada por ganado, la mayoría de las especies desaparecerá, y sólo permanecerán aquellos organismos cuyas características abrasivas le proporcionan resistencia ante el pastoreo.
Es necesario reiterar que todos los tipos de depredación son interacciones en las que la selección natural tiene un papel importantísimo en la regulación de las relaciones entre las especies (véase el tema Evolución); es por eso que a lo largo de la evolución, tanto depredadores como presas han desarrollado diversos mecanismos que les permiten adaptarse a las condiciones cambiantes del ambiente.
Este es un tipo de interacción en la que ambas especies reciben beneficios, mejorando con ello su desempeño individual. Existen diversos grados de asociación, de tal manera que es posible distinguir a los mutualistas obligados y los facultativos. Los mutualistas obligados son especies que dependen de manera total una de la otra para su supervivencia; por ejemplo, la asociación entre las termitas del orden isóptera y el protista flagelado Trichomonas termopsidis, que habita en su tracto digestivo. La termita le brinda al protista protección y alimento; por su parte, el protista fermenta la celulosa proveniente de la madera y deja escapar dióxido de carbono e hidrógeno; el ácido acético, que es el principal producto de la fermentación, es absorbido por la termita a través de su intestino. Esta dependencia es tal que si se separaran ninguno de los dos organismos sobreviviría.
En el rumen (una de las cavidades del aparato digestivo de las vacas y otros bóvidos) existe una compleja comunidad de microorganismos constituida por bacterias anaerobias y protistas, muchos de ellos especializados, que en conjunto tienen la capacidad de digerir la celulosa y otros componentes de los tejidos vegetales que los rumiantes por sí solos no podrían digerir; por su parte, la microflora del rumen tiene garantizado un aporte continuo de alimento y un lugar donde vivir.
El mutualismo intracelular se ha documentado en ciertas especies de insectos como los áfidos y las cucarachas. En estos casos, los huéspedes intracelulares son ciertas bacterias que tienen la capacidad de sintetizar algunos aminoácidos indispensables para los insectos. Sin embargo, el mutualismo intracelular no se limita a los insectos, ya que también se halla entre protistas y bacterias; por ejemplo, el protista Paramecium bursaria alberga en su interior una gran cantidad de cianobacterias (bacterias fotosintetizadoras), que proporcionan al huésped compuestos de carbono fabricados por ellas, mientras el protista les brinda materias primas para la elaboración del alimento y hábitat. Existe una fuerte evidencia que indica que los mutualismos intracelulares jugaron un papel central en una de las etapas cruciales de la evolución, que dio origen a mitocondrias y cloroplastos de las células eucariontes (véase el tema Biología celular, molecular y bioquímica).
Otro caso bien documentado es la asociación que existe entre un arbusto tropical (Acacia cornigera) y las hormigas del género Pseudomyrmex. Las hormigas viven en el interior de las espinas huecas de la planta y se alimentan con un producto azucarado que ésta produce. A cambio, las hormigas mantienen alejadas a otras especies de insectos que pudieran ser depredadores potenciales de las acacias. Se han hecho experimentos en los que colonias completas de hormigas se removieron de varios arbustos, y se observó que al poco tiempo éstos fueron presa de otras especies de insectos fitófagos que les causaron daños severos.
Es necesario resaltar que el mutualismo no es una relación de tipo altruista, ya que ninguno de los organismos actúa en beneficio del otro, sino para el propio. Sin embargo, como resultado de la asociación ambas especies se benefician. En la mayoría de las relaciones mutualistas, una de las especies le brinda alimento a la otra, mientras esta última le proporciona a la primera limpieza, protección, dispersión, polinización o un lugar donde vivir.
Existen otros casos interesantes de relaciones mutualistas. Uno de los más extendidos es la dependencia que existe entre las plantas con flores y los polinizadores. Las plantas desarrollaron estrategias para atraer a polinizadores específicos, quienes, por su parte, han perfeccionado mecanismos para el reconocimiento de plantas específicas. Por ejemplo, el mirto rojo (Salvia fulgens) es polinizado por el colibrí, mientras que la polinización de las dos especies de maguey de interés comercial —el pulquero (Agave salmiana) y el tequilero (Agave tequilana)— depende de dos especies de murciélagos del género Leptonycteris.
Muchos polinizadores pueden asociarse con varias especies de plantas. Sin embargo, existen algunas familias, como las orquídeas, en donde se observa un grado alto de especialización de las estructuras florales que restringen el acceso a unas pocas especies de insectos.
Aparte de la polinización, algunas relaciones mutualistas propician la dispersión de semillas, como ocurre con varias especies de aves, roedores, murciélagos y monos, que al alimentarse de los frutos de ciertas plantas esparcen las semillas lejos de las plantas progenitoras, a través de los residuos que caen al suelo o por medio de las heces fecales. Existen otros ejemplos de mutualismo, en los que una de las especies recibe servicios de limpieza o protección en contra de depredadores, a cambio de alimento o un lugar donde vivir. Es el caso de los peces payaso que habitan entre los tentáculos de las anémonas, o los peces y pequeños crustáceos marinos que mantienen limpia la piel o agallas de otros peces al consumir sus parásitos.
En los mutualismos muy especializados se da una dependencia extrema por parte de las especies que participan en ella, a tal grado que existe pérdida de la individualidad, pues algunas de las funciones vitales no pueden ser llevadas a cabo de manera independiente por las especies involucradas, como es el caso de la asociación que existe entre los hongos y algas que forman el liquen o la que existe entre la flora intestinal y el rumen de los bóvidos. Estos casos son un claro ejemplo de coevolución.
Es un tipo de relación en que una de las especies recibe beneficios, mientras la otra no es afectada. Una asociación de este tipo es la que se da entre diversas plantas epífitas (plantas que viven sobre otras plantas), como las bromelias y orquídeas, y ciertas especies de árboles en los bosques tropicales. No obstante, si la cantidad de epífitas es grande, pueden llegar a causar ciertos efectos negativos en los árboles en los que se encuentran.
Se caracteriza por ser una interacción en la que una especie provoca daños en otras, pero éstas no tienen ningún efecto bueno o malo sobre la primera. Un ejemplo es la producción de sustancias por parte de las hojas del eucalipto, que inhiben el crecimiento de otras plantas en las zonas donde caen las hojas de este árbol. A este fenómeno se lo denomina alelopatía.
Hasta ahora se han revisado las características de las interacciones ecológicas en las que intervienen unas cuantas especies. Sin embargo, como se ha visto a lo largo de este tema, las complejas interacciones que se dan entre los miembros integrantes de las comunidades pueden tener efectos directos sobre las especies involucradas o bien indirectos, lo que causa alteraciones en la dinámica de las poblaciones.
La diversidad de interacciones que se presenta en las comunidades tiene un papel fundamental en la estabilidad, así como en la capacidad homeostática de las mismas. En este sentido, actualmente hay una tendencia a estudiar las interacciones en el contexto integral de la comunidad, por lo que los modelos de la interacción de dos especies sirven únicamente como marco de referencia.
